
Apabila ada pertanyaan tentang contoh hubungan mutualisme di alam, jawaban rayap dan flagellata, yakni suatu protista berflagel dalam perut rayap, biasanya akan muncul. Namun, simbiosis mutualisme rayap ternyata bukan hanya dengan flagelata, melainkan dengan segenap isi perutnya. Tidak hanya rayap, ternyata kecoak juga menjalin hubungan simbiosis mutualisme dengan mikroba penghuni badannya.
Pertama-tama, mari kita telaah klasifikasi rayap. Dulu rayap ditempatkan dalam ordo tersendiri, yaitu Isoptera (iso: sama; ptera: sayap). Namun, berdasarkan bukti molekuler, rayap ternyata berkerabat dekat dengan kecoak. Mereka pun dikumpulkan dalam ordo Blattodea.
Anggota ordo Blattodea menunjukkan ciri berupa perilaku coprophagy (memakan kotoran). Perilaku tersebut tidak hanya bertujuan untuk mendapatkan nutrien, tetapi juga untuk transfer isi perut, yang mengandung ribuan jenis mikroba. Organisme mikroskopis ini sangat penting untuk kelangsungan hidup anggota Blattodea. Simbiosis antara Blattodea dan mikroba simbionnya berdasarkan premis inang rayap atau kecoak mendapatkan nutrien tambahan dari simbion, sedangkan simbion mendapat tempat hidup aman dan asupan nutrien yang teratur dibandingkan ketika harus hidup di alam liar.
Hampir semua spesies kecoak memiliki bakteri endosimbion obligat, Blattabacterium, yang hidup di bagian fat body (jaringan penyimpan lemak). Istilah obligat berarti baik kecoak maupun bakteri endosimbion tersebut tidak bisa hidup sendiri tanpa lainnya. Uniknya, Blattabacterium ternyata juga ditemukan pada spesies rayap Mastotermes darwiniensis, simbion rayap yang biasanya ditemukan dalam usus perut, bukan di dalam jaringan tubuh. Hadirnya Blattabacterium pada spesies rayap tersebut membuktikan bahwa pada suatu waktu rayap dan kecoak berasal dari nenek moyang yang sama.


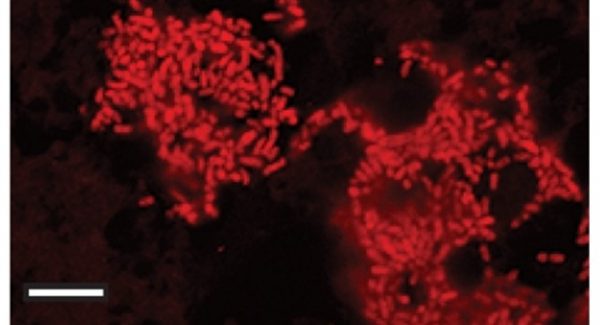
Blattabacterium memiliki fungsi vital dalam siklus nitrogen pada tubuh kecoak. Nitrogen merupakan unsur penting pembentuk protein, sedangkan makanan kebanyakan jenis kecoak hanya mengandung sedikit nitrogen. Dengan adanya bakteri simbion ini, asam urat (uric acid) yang biasanya dibuang, setelah dipecah oleh mikroba usus menjadi urea dan/atau amonia dapat didaur ulang oleh Blattabacterium menjadi asam amino. Bayangkan apabila manusia memiliki kemampuan itu, urea dalam urin kita bisa diubah kembali menjadi protein, yang artinya kita hanya perlu makan sumber protein dari luar dengan sekadarnya. Itulah mengapa kecoak sangat tangguh. Mereka hanya butuh sedikit makanan untuk bertahan hidup, karena bakteri endosimbion dalam tubuhnya mengoptimalkan pemakaian nutrien.
Pada rayap, mikroba simbion hidup dalam usus bukan dalam jaringan tubuh (selain M. darwiniensis yang memiliki keduanya). Simbion rayap tersebut terdiri dari jenis eukariot (protista) dan prokariot (bakteria dan arkaea) yang mayoritas hidup sebagai endosimbion atau ektosimbion pada protista penghuni usus. Usus rayap merupakan tempat yang menakjubkan. Dalam tempat sekecil itu terdapat puluhan ribu sel protista dan ratusan juta sel prokariot. Sayangnya, sebagian besar dari mereka tidak dapat dikultivasi.
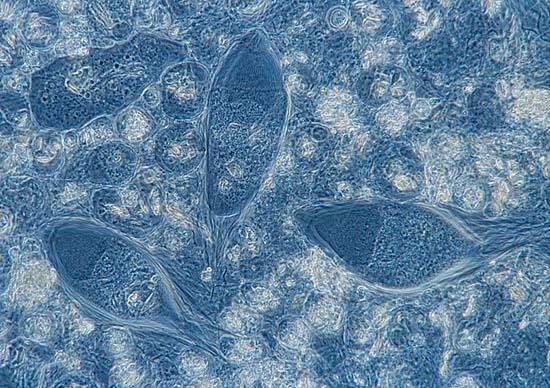
Sama seperti kecoak, makanan utama rayap adalah tumbuhan yang sudah membusuk, bahkan ada juga yang bisa hidup dari kayu kering atau lembaran kertas yang tentu saja hampir tidak mengandung nitrogen. Kertas tersusun dari selulosa, yang sejatinya hanya mengandung unsur karbon, oksigen dan hidrogen. Bagaimana rayap bisa bertahan hidup dengan makanan yang hanya sedikit atau tanpa unsur nitrogen? Di sinilah peran simbion usus rayap.
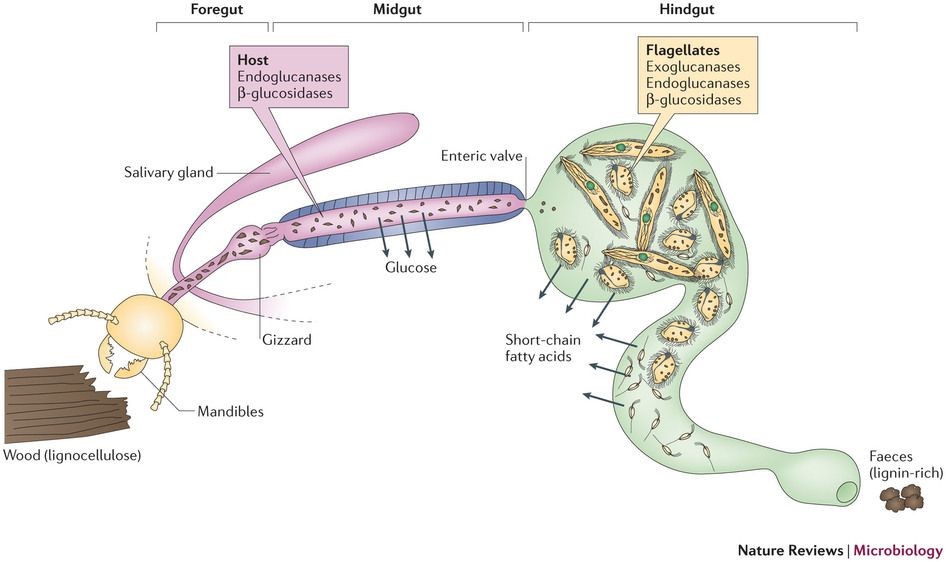
Kayu, kertas dan makanan lain yang dimakan rayap, setelah dikunyah menjadi bentuk yang lebih kecil, diurai menjadi bentuk yang lebih sederhana. Protista dalam usus rayap sangat berperan dalam tahap ini. Protista, yang kebanyakan termasuk dalam flagellata, merombak dan memfermentasi hampir secara total partikel kayu yang bersebaran dalam usus menjadi asetat, H2 dan CO2.
Asetat dapat digunakan oleh rayap sebagai sumber energi dan karbon, sedangkan H2 dan CO2 dapat dimetabolisme menjadi asetat oleh bakteri usus melalui asetogenesis reduktif (reductive acetogenesis). Metabolisme asetogenesis reduktif ini sangat spesial karena asetat dihasilkan hanya dengan hidrogen dan karbondioksida. Biasanya asetat dihasilkan dari reaksi kimia yang lebih panjang. Selain itu, bakteri usus rayap juga mampu mendaur ulang sampah nitrogen dari rayap dan protista, dan menambat nitrogen dari udara menjadi amonia kemudian mengubahnya menjadi asam amino sehingga dapat menutupi kebutuhan nitrogen inang rayap.
Rayap dan simbionnya telah berevolusi (co-evolved) dan berspesiasi (co-speciation) bersama selama jutaan tahun. Oleh karena itu, protista dan beberapa bakteri endosimbion yang dijumpai dalam usus suatu jenis rayap, hanya spesifik ditemukan di jenis rayap tersebut, tapi tidak di jenis lain. Misalnya, protista dari genus Pseudotrichonympha hanya ditemukan pada rayap famili Rhinotermitidae, kecuali pada genus Reticulitermes. Di dalam sitosol Pseudotrichonympha ditemukan bakteri endosimbion Azobacteroides yang dapat menambat nitrogen (diazotrof). Bakteri diazotrof dan inang Pseudotrichonympha ini tidak memiliki kerabat dekat yang hidup di alam bebas karena mereka sudah begitu terspesiasi selama jutaan tahun di dalam usus rayap.
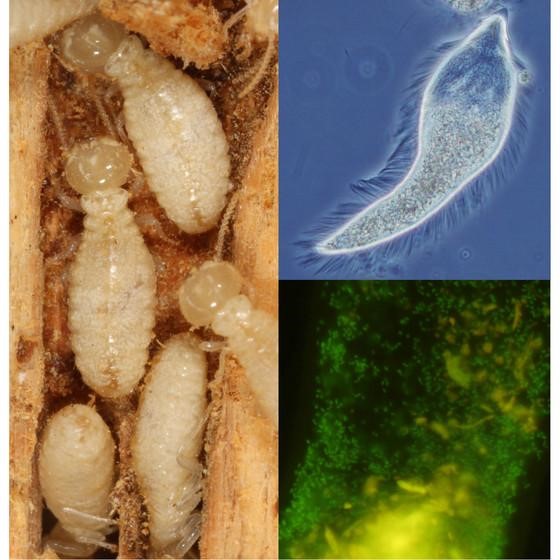
Ada pula protista dari genus Trichonympha, yang menghuni usus pada berbagai jenis rayap dan kecoak. Trichonympha menjadi inang untuk bakteri endosimbion (dari bakteri kelas Endomicrobia) dalam sitosolnya dan ektosimbion (spirochetes) yang menempel pada permukaan selnya. Ada pula bakteri endosimbion yang hidup di dalam inti sel sebagaimana yang ditunjukkan oleh Verrucomicrobia yang hidup dalam nukleoplasma Trichonympha agilis. Bakteri endosimbion dalam nukleoplasma juga ditemukan pada protista usus rayap lain, misalnya pada Pyrsonympha grandis, Dinenympha exilis, Dinenympha parva dan Teranympha mirabilis.
Setelah membaca uraian di atas, semoga sekarang kita bisa menjawab pertanyaan “Apa contoh simbiosis mutualisme di alam?” dengan lebih lengkap. Tidak cukup dengan rayap dan flagelata, tetapi lebih tepatnya rayap dan isi perutnya, dan juga bisa ditambahkan dengan kecoak dan endosimbionnya.
Bahan bacaan:
- Brune, Andreas. (2014). Symbiotic digestion of lignocellulose in termite guts. Nature Reviews Microbiology. https://doi.org/10.1038/nrmicro3182
- Hongoh, Y. dkk. (2008). Genome of an endosymbiont coupling N2 fixation to cellulolysis within protist cells in termite gut. Science 322, 1108–1109.
- Latorre, A. 2015. https://www.slideshare.net/FundacionAreces/amparo-latorre-simposio-microbiologa-transmisin
- Kuwahara, H. dkk. (2016). Genome of ‘Ca. Desulfovibrio trichonymphae’, an H2-oxidizing bacterium in a tripartite symbiotic system within a protist cell in the termite gut. The ISME journal, 11(3), 766–776. doi:10.1038/ismej.2016.143
- Sato, T. dkk. (2013). Intranuclear verrucomicrobial symbionts and evidence of lateral gene transfer to the host protist in the termite gut. The ISME journal, 8(5), 1008–1019. doi:10.1038/ismej.2013.222Sabree, Z. L., Kambhampati, S., & Moran, N. A. (2009). Nitrogen recycling and nutritional provisioning by Blattabacterium, the cockroach endosymbiont. Proceedings of the National Academy of Sciences of the United States of America, 106(46), 19521–19526. doi:10.1073/pnas.0907504106
- Van Egmond, W. 2004. http://www.microscopy-uk.org.uk/mag/wimsmall/termite.html
Penulis:
Ajeng Kusumaningtyas Pramono, alumnus Tokyo Institute of Technology, Jepang, pernah meneliti gut microbiome. Kontak: ajengpramono(at)gmail(dot)com


