
Reproduksi pada tumbuhan dapat terjadi baik secara seksual maupun aseksual. Reproduksi seksual pada tumbuhan membutuhkan pembuahan sel telur (ovum) oleh serbuk sari (polen). Pembuahan ini dapat terjadi melalui sistem perkawinan sendiri (selfing) ataupun perkawinan silang (outcrossing).
Sistem perkawinan tumbuhan dipengaruhi oleh banyak faktor. Salah satunya adalah arsitektur bunga dan pembungaan. Secara umum, bunga dapat dikategorikan sebagai bunga hermafrodit dan dioecy. Bunga hermafrodit merupakan bunga sempurna yang memiliki putik dan benang sari. Namun, meskipun bunga hermafrodit memiliki organ seksual yang lengkap, variasi masih dapat terjadi dan mampu memengaruhi sistem perkawinan pada tumbuhan. Variasi tersebut dapat berupa polimorfisme pengaturan posisi organ seksual.
Heterostili merupakan bentuk polimorfisme ketika suatu jenis tumbuhan memiliki perbedaan panjang relatif antara tangkai putik (style) terhadap benang sari (stamen) pada bunga dari individu yang berbeda. Populasi heterostili terdiri atas dua (distyly) atau tiga (tristyly) tipe morfologi bunga yang dapat dibedakan dari posisi resiprok kepala putik (stigma) dan kepala sari (anther). Fenomena ini pertama kali dikemukakan oleh Charles Darwin dalam tulisannya yang berjudul The Different Forms of Flowers on Plants of the Same Species.
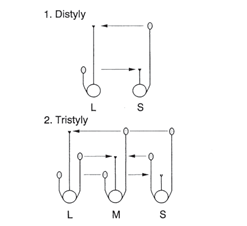
Heterostili ditemukan terjadi pada sekitar 28 marga tumbuhan Angiospermae dan diyakini merupakan perangkat tumbuhan untuk melakukan perkawinan silang dengan perantara hewan sebagai polinator. Adanya perbedaan posisi stigma dan anther pada tumbuhan serta heterostili adalah strategi tumbuhan untuk mengurangi pembuahan sendiri atau dikenal sebagai resiprokal herkogami.
Darwin dalam pengamatannya menyadari bahwa morfologi yang resiprok membuat tumbuhan menempatkan polen pada bagian tubuh yang berbeda dari polinator dan mampu memfasilitasi polinasi antartipe bunga yang berbeda. Penelitian-penelitian selanjutnya mendukung hipotesis Darwin tersebut dan menunjukkan bahwa heterostili mampu memfasilitasi terjadinya perkawinan silang.
Fenomena heterostili telah mengundang banyak pertanyaan dari segi evolusi, ekologi, maupun genetik. Hal ini menjadikan heterostili banyak diteliti dan merupakan salah satu bentuk adaptasi tumbuhan yang paling dipahami karena banyaknya penelitian mengenai fenomena ini. Secara evolusi telah diketahui bahwa heterostili berkembang secara independen pada berbagai marga tumbuhan dan tidak saling terkait antara satu marga dan marga lainnya (multiple independent origin).
Terdapat dua model teori asal usul distylyI, yaitu selfing avoidance model yang dikemukakan oleh Charlesworth dan pollen transfer model yang dikemukakan oleh Lloyd & Webb. Kedua model ini berbeda pada fenotipe dari populasi tetua. Model asal-usul tristyly hanya dikemukakan oleh Charlesworth yang melibatkan parameter transfer polen, selfing, dan inbreeding depression. Penjelasan yang komprehensif mengenai evolusi distyly dan tristyly dikemukakan oleh Barrett dengan visualisasi yang ditunjukkan pada gambar.
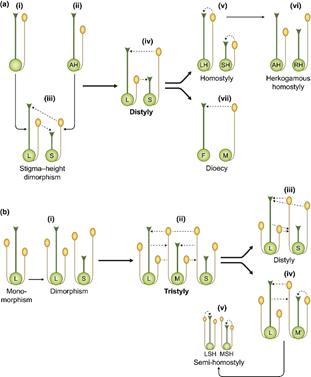
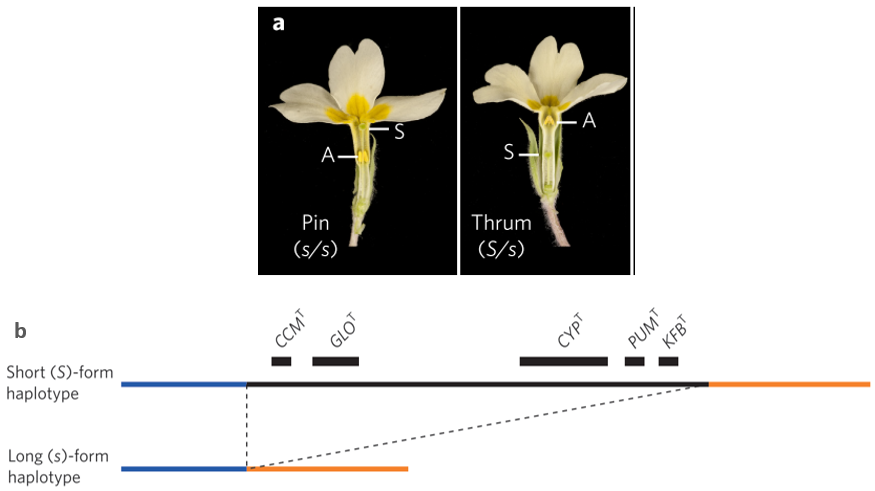
Variasi panjang style telah lama dikaji secara genetik dan diketahui diatur oleh kompleks S-locus supergene. Genotipe heterostili dijelaskan dengan baik pada populasi tumbuhan Primula yang memiliki dua tipe bunga, yaitu pin dan thrum. Individu tumbuhan yang memiliki tipe bunga dengan style pendek (thrum/S-morph) mempunyai genotipe heterozigot S/s, sementara individu yang memiliki tipe bunga dengan style panjang (Pin/L-morph) mempunyai genotipe homozigot s/s.
Adapun pada populasi tristyly dijelaskan bahwa alel dominan pada S-locus menyebabkan tipe bunga S-morph, sementara individu dengan homozigot resesif s/s dapat memiliki fenotipe bunga L-morph ataupun M-morph tergantung genotipe pada lokus M. Individu tumbuhan dengan genotipe s/s M/m dan s/s M/M akan memiliki fenotipe bunga M-morph (mid length style) dan individu dengan genotipe s/s m/m memiliki genotipe L-morph.
Analisis arsitektur genetik S-locus supergene pada tumbuhan P. vulgaris menuntun pada pengetahuan bahwa S-locus supergene adalah 5 klaster gen spesifik thrum yang berkaitan erat (linked cluster genes) berukuran 278 kb. Lima klaster gen tersebut adalah CCMT, GLOT, CYPT, PUMT, dan KFBT. Klaster-klaster gen ini ditemukan pada individu tumbuhan dengan fenotipe bunga thrum dan tidak ditemukan pada tumbuhan dengan bunga pin. Apabila dihubungkan dengan pengetahuan sebelumnya, dapat disimpulkan bahwa kelima klaster gen tersebut adalah alel dominan dari S-locus yang ekspresinya menyebabkan terbentuknya fenotipe bunga S-morph.
Bahan Bacaan:
- Barrett, S.C.H., 2019. ‘A most complex marriage arrangement’: recent advances on heterostili and unresolved questions. New Phytol. 224: 1051–1067. https://doi.org/10.1111/nph.16026
- Barrett, S.C.H., 1992. Heterostylous genetic polymorphisms: model systems for evolutionary analysis, In: Barrett, S.C.H. (Ed.). Evolution and Function of Heterostili. Springer-Verlag Berlin Heidelberg. New York.
- Kappel, C., Huu, C.N., Lenhard, M., 2017. A short story gets longer : recent insights into the molecular basis of heterostili. J. Exp. Bot. 68: 5719–5730. https://doi.org/10.1093/jxb/erx387
- Li, J., Cocker, J.M., Wright, J., Webster, M.A., Mcmullan, M., Dyer, S., Swarbreck, D., Caccamo, M., Oosterhout, C. Van, Gilmartin, P.M., 2016. Genetic architecture and evolution of the S locus supergene in Primula vulgaris. Nat. Plants 2: 1–7. https://doi.org/10.1038/nplants.2016.188
- Mcclure, B., 2016. The genetic basis of heterostili. Nat. Plants 2: 1–2. https://doi.org/10.1038/NPLANTS.2016.184
Penulis:
Seni Kurnia Senjaya, Peneliti di Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia.
Kontak: senikurnia(at)gmail.com


